|
|
|
   |
הסדרי נגישות
|
| עמוד הבית > מדעים > אקולוגיה ואיכות הסביבה > חורש ים-תיכוניעמוד הבית > ישראל (חדש) > אקולוגיה ואיכות הסביבה > שטחים פתוחיםעמוד הבית > ישראל (חדש) > נוף וטבע > צמחים |
|
|||||||||||||||||||||||||||||||||||||||||||||
|
|
המאפיינים האקלימיים של האזורים הים-תיכוניים (ראה פרק האקלים הים-תיכוני), מביאים להיווצרות תנאים של עקת (stress) יובש וחום במשך הקיץ הארוך יחסית. חלק מכריע של תכונות ההתאמה המיוחסות לצמחי החורש הים-תיכוני מוסבר לפיכך כעמידות ליובש ולחום, דהיינו: הימנעות מאיבוד יתר של מים ויכולת לאזן את משק המים באופן שישמר כושר לקיום פוטוסינתזה, גם בתנאים של צמצום באספקת המים ובטמפרטורה גבוהה. הדמיון לצמחי מדבר מתבקש מאליו, אולם יש להדגיש, שבניגוד למערכת המדברית, שהיא הרבה יותר קיצונית מבחינת מיעוט המים ומאופיינת בוואריאביליות גבוהה ויכולת חיזוי נמוכה של משק המים - המערכת הים-תיכונית היא מערכת חזויה וודאית במידה רבה יחסית. למרות שיש תנודות בכמות המשקעים משנה לשנה, הרי חורף גשום, במידה זו או אחרת, הוא נתון קבוע בכל האזורים הים-תיכוניים בעולם. לפיכך, תכונות ההתאמה של צמחי החורש ליובש ולחום הקיץ צריכות להיבחן תוך ידיעה שהמים בכל זאת זמינים בחלק נכבד של השנה. טמפרטורות החורף באזורים ים-תיכוניים אמנם אינן יורדות, בדרך כלל, לערכים נמוכים מדי (ראה פרק האקלים), אולם ייתכן שלעתים גם טמפרטורות החורף עלולות להוות גורם מגביל לתהליכים שונים. מלבד המכלול האקלימי המיוחד לסובב הים-תיכוני, הקרקעות של האזורים הים-תיכוניים מאופיינות בדלות יחסית של מזינים מינרליים, ובעיקר חנקן וזרחן. גם זו עקה סביבתית שיש להביאה בחשבון כאשר דנים בהתאמות צמחי החורש. להלן נסקור התאמות של צמחי חורש לתנאי הסובב הים-תיכוני בכמה היבטים: פנולוגיה, מאזן המים, פוטוסינתזה, פרודוקטיביות, ופעילות קמביאלית (יצירת תאים חדשים של צינורות הובלה בצמח ע"י רקמת הקמביום, שתוצאתה – צמיחה לרוחב והתעבות של גבעולים או שורשים). צמחי החורש הים-תיכוני מושפעים גם מבעלי החיים הנמצאים במערכת, הן הטבעיים והן אלה שהאדם ביית. סביר מאוד שבצמחי החורש מופיעות תכונות התאמה גם למרכיבים אלה של המערכת האקולוגית. נייחד לפיכך מקום גם למאפיינים מיוחדים של האבקה והפצת פירות וזרעים בצמחי החורש הים-תיכוני.
במגוון צורות החיים של החורש הים-תיכוני, אין ספק שהדומיננטיים הם העצים והשיחים, ולכן נעסוק להלן בעיקר בתכונותיהם של אלה. באופן גס וכוללני, אפשר להבחין בהם ב-3 קבוצות, על יסוד המורפולוגיה של העלים:
הצמחים הסקלרופיליים ירוקי-העד והצמחים משירי העלים בקיץ הם הדומיננטיים בכל התצורות הים-תיכוניות, והם גם המזהים האופייניים ביותר שלהן. הטיפוסים האחרים מלווים אותם, אך אינם מצויים בכל מקום. למשל, הטיפוס ירוק-העד צר העלים מופיע בקליפורניה ובדרום אפריקה, אך נעדר מאגן הים התיכון. הצמחים המאלאקופיליים נשירי החורף שכיחים יחסית במזרח אגן הים התיכון, כנראה בשל חדירת צומח מאסיבית, בתקופות אקלימיות שונות, מהאזור הערבתי, אירנוטורני. הם נפוצים יותר בחגורות רום גבוהות יותר באזורים הים-תיכוניים, בעלות אקלים יותר קר וגשום אך יש להם נציגות גם באזורים "רגילים" (לדוגמה: אלה ארץ ישראלית). נשירי חורף אינם גדלים בצ'אפארל של קליפורניה וגם באזורי החורש הטיפוסיים של מערב הים התיכון הם נדירים. מעניין שדווקא במזרח הים התיכון חסרים כמעט לחלוטין נשירי הקיץ, שאמורים להיות בעלי התאמה טובה לעקת היובש. ייתכן והדבר קשור גם לעובדה שמרבית בתי הגידול בנופים אלה עניים במינרלים, ואחת ההתאמות לבית גידול עני היא השארת עלים לאורך זמן, ובכך הקטנת השימוש במינרלים חסרים או ויסות השימוש בהם לאורך תקופת זמן ארוכה יחסית (1977 ,Grime). מעניין לציין שניתוח מקיף של צומח באזור שאינו ים-תיכוני (מרכז פלורידה) הראה קשר ברור בין צמחים ירוקי-עד ובין אתרים יבשים ועניים במינרלים (Monk, 1966). הטיפוסים האקומורפולוגיים השונים של צמחי החורש מבטאים הן התאמה לקיום במיקרו-בתי גידול שונים והן דרכים שונות לניצול משאבי הסביבה. בהכללה, ניתן לומר שהצמחים הלא-סקלרופיליים, משירי עלים בקיץ, גדלים בבתי גידול יותר יובשניים לעומת הצמחים הסקלרופיליים ירוקי העד. בתי גידול יותר יובשניים משמעם מפנים דרומיים, אזורים פחות גשומים או קרקעות רדודות וסחופות. מאידך גיסא, צמחים מאלאקופיליים ירוקי-עד, כגון ער אציל, גדלים במיקרו בתי גידול לחים יותר - כמו אתרים יותר גשומים או במדרונות צפוניים (1973 ,Walter ;1947 ,(Boyko. בהקשר זה מעניינת ההשוואה שעשו מוני ודון (1970 ,Mooney + Dunn) בין מינים ירוקי-עד ונשירי קיץ, המופיעים בקליפורניה וצ'ילי. הם מצאו ששיחים נשירי קיץ מחליפים עצים ירוקי-עד כאשר היובשנות עולה. לירוקי העד ככלל יש מערכת שורשים מפותחת יותר, עונת ההטמעה ארוכה יותר אך שיעור ההטמעה נמוך יותר. לנשירי הקיץ, לעומת זאת, יש שיעור קיבוע פחמן גבוה, אבל לאורך תקופה קצרה יותר, וגם מערכת השורשים רדודה יותר. במילים אחרות, ירוקי העד משקיעים משאבים כדי לפתח סובלנות ליובש המאפשרת ייצור לאורך השנה כולה. ככל שהתנאים יבשים יותר פוטנציאל ההטמעה של ירוקי העד יורד, ואז אין להם יתרון על נשירי הקיץ. עצים נשירי חורף, כמו מינים של צפצפה, דולב, אולמוס ואחרים, משתלבים בחורש בתנאים של משק מים משופר מאוד, בעיקר בקרבת נהרות או נחלים (1973 ,Walter). מכיוון שהצמחים הסקלרופיליים והצמחים משירי העלים בקיץ שאינם סקלרופיליים הם המבטאים המהימנים של התאמה לתנאים ים-תיכוניים, יוקדש להם עיקר הדיון שלהלן. אולם, קודם לכן, יש צורך בהגדרות מדויקות יותר של הסקלרופיליות ובהכרת המאפיינים שלה.
הסקלרופיליות של צמחים ירוקי-עד ניתנת לאיפיון בעיקר לפי תכונות העלים. לפי Margaris (שנת 1981) Miller (שנת 1983), עלים של צמחים סקלרופיליים הם בדרך כלל קטנים, עבים וגלדניים. עם זאת, הם יכולים להיות נבדלים ביניהם בגודל. Christodulakis ו-Mitrakos (שנת 1987) חילקו את העלים של הצמחים הסקלרופיליים השונים ל-4 קבוצות גודל על-פי שטח העלה במילימטרים רבועים, כדלקמן:
Grime (שנת 1997) טוען שצמחים סקלרופיליים הם הנפוצים ביותר באזורים בהם העונה הרטובה קצרה. לפי Walter (שנת 1973), היתרון בסקלרופיליות הוא היכולת להמשיך ולנהל חילופי גזים כאשר יש מספיק מים בצמח, אך להוריד דרסטית את חילופי הגזים, בעזרת סגירת פיוניות, כאשר המים חסרים. לפי Mooney וחובריו (1974, מצוטט אצל Miller, 1983) יש קשר בין גודל העלים וצורתם לבין מידת היובשניות בבית הגידול. ככל שהיובשניות בצ'אפארל הולכת וגדלה, העלים נעשים צרים יותר ונטויים. מידת הסקלרופיליות של העלים ניתנת לאיפיון בעזרת מדדים שונים. לפי קומרוב (1973 ,Kummerow), עלים סקלרופיליים מאופיינים ביחס נמוך של שטח פנים לנפח. Margaris (שנת 1981), מציין שעלים של צמחים סקלרופיליים מצטיינים במסה גבוהה ליחידת שטח, כלומר, בצפיפות גבוהה. הוא קובע שהצפיפות הגבוהה של העלה בצמח ירוק-עד חשובה, לעלה בעל משך חיים ארוך, לשמירה על מאזן המים ולהגנה מפני טורפים. מצד שני מבנה כזה מקטין את הכושר לחילוף גזים. קריסטודולאקיס ומיטראקוס (1987) סיכמו נתונים על היחס בין משקל העלים לבין שטח הפנים שלהם ועל צפיפות העלה בעצים ושיחים נפוצים של חורש ים-תיכוני ביוון, שחלקם גדלים גם בארץ (טבלה 5).  טבלה 5 מראה, שלמרות שההתאמה בין שני המדדים אינה תמיד מלאה, הרי בדרך כלל מדדי הסקלרופיליות גבוהים יותר בצמחי חורש ירוקי-עד מאשר בכמה צמחי תרבות או עצי-בר נשירי חורף. מעניינת העובדה עד כמה נמוכים ערכי הסקלרופיליות שהתקבלו עבור ער אציל, התואמים את שיוכו של מין זה ע"י וולטר לקבוצת המינים המאלאקופיליים (ראה לעיל). גם Eherlinger ו-Mooney (שנת 1983) ו-Harley וחובריו (1987) מציינים שמשקל סגולי גבוה של עלים הוא תכונה המאפיינת עלים סקלרומורפיים, בהשוואה לעלים של צמחים ים-תיכוניים נשירי קיץ. סקלרופיליות מבטאת איפוא, השקעה רבה של חומר יבש והקצאה מוגברת של משאבי פחמן לבניין העלה. השקעה זו מקנה חוזק מכני לעלים ארוכי חיים בצמחים ירוקי-עד, והיא נובעת מהרבדה מאסיבית של תאית, ליגנין וחומרים משניים בתאים (1983 ,Miller). בעקבות זאת, מוסט שיווי המשקל ביחס פחמן:חנקן בעלים לטובת הפחמן. עלים כאלה הם "יקרים" יותר לייצור בהשוואה לעלים של צמחים משירי עלים. ה"פיצוי" או החזרת ההשקעה הוא במשך הזמן הארוך של יכולת פעילות פוטוסינתטית. לפי קומרוב, מהווה הסקלרופיליות ביטוי לגירעון במים, עוצמת אור גבוהה ומחסור בחנקן בבית הגידול. החורש מתפתח באזורים המאופיינים ברמת חנקן נמוכה מחד גיסא, ומאידך גיסא, בתנאי אקלים ונוף המאפשרים הטמעה רצופה לאורך זמן רב והיוצרים "עודפי" פחמן בצמח. עובדה זו מאפשרת "השקעה" מסיבית בכיוון המבני - גלדניות, אורך חיי עלה. עדות לכך שמשקל סגולי גבוה יותר (=סקלרופיליות מוגברת) קשור לעמידות ליובש מובאת ע"י רמבל וליטם (1987 ,Leteme + Rambal). הם מצאו באלון קוקציפרה (קרוב מאוד לאלון מצוי בארץ) עלייה גדולה במשקל הסגולי של העלים לאורך מפל גיאוגרפי מדרום צרפת לספרד, שבמהלכו כמות המשקעים השנתית יורדת מ-1188 עד ל-301 מ"מ. העלים הסקלרומורפיים נישאים, כאמור, על צמחים ירוקי-עד. Mooney (שנת 1983), מציין שאורך החיים של העלים הללו מגיע עד 3 שנים. למרות שעונת הלבלוב מוגבלת בדרך כלל לאביב, הצמח נושא תמיד עלים ירוקים בעלי כושר פוטוסינתטי. Pereira וחובריו (1987) מצאו במחקריהם בפורטוגל שאורך החיים הממוצע של עלים במינים סקלרופיליים הוא פחות משנה. באלון קוקציפרה, באלון השעם ובקטלב אונדו, רוב העלים שנוצרים בגל הליבלוב האביבי נושרים תוך 13 חודש, ומיעוטם ממשיך עד לגיל של 17 חודש. היותם של העצים ירוקי-עד מתאפשר הודות לאותו חלק של העלווה שנשאר על הענפים למעלה משנה ומעלים חדשים של ליבלוב צעיר שמתפתחים בצורה מתוזמנת עם השרת חלק מהעלים הקודמים. אם משווים לכך את ההתנהגות של עלים בצמחים ירוקי-עד לא-סקלרופיליים, שמשירים חלק מעליהם בקיץ, מתקבלת תמונה שונה. Pereira וחובריו (1987) מצאו שבמין הלוטם (salvifolius Cistus) אורך החיים הממוצע של העלים הוא 131 יום בלבד. חלק גדול מהעלים שליבלבו בחורף נושר בקיץ, ואילו העלים ששורדים בקיץ ומאפשרים לצמח לקיים פוטוסינתזה, הם עלים צעירים שליבלבו באביב המאוחר או בראשית הקיץ. סביר להניח, שיש קשר בין ההשקעה הרבה יחסית בייצור עלה סקלרופילי לעומת ההשקעה בעלה הלא-סקלרופילי לבין אורך חייהם השונה. הבדלים אלה בין משך החיים של עלים סקלרופיליים לבין אלה של עלים לא-סקלרופיליים צמודים להבדלים בפנולוגיה (ראה להלן). ככלל, עונת חידוש העלווה אצל הסקלרופיליים היא באביב, ונמשכת זמן קצר יחסית. לעומת זאת, בצמחים לא-סקלרופיליים, כמו בלוטם, הצמיחה משתרעת על כל חודשי החורף והאביב עד לקיץ המוקדם. הסקלרופיליים, בעלי עלים ארוכי חיים יחסית, מנצלים, לצימוח מחודש במשך זמן קצר, משאבים ומלאים שנוצרו קודם לכן ע"י העלווה הירוקה. הלא-סקלרופיליים, מנצלים את התוצרים של ההטמעה השוטפת. תופעה דומה נצפתה גם באלון מצוי ומינים אחרים של החורש הים-תיכוני בארץ (אורשן, 1989).
השאלה המרכזית לגבי התאמתם של צמחי החורש הים-תיכוני לתנאי הסובב היא יכולתם להתמודד עם היובש והחום של הקיץ. התאמות אלה נבחנות בכמה רמות:
כל הרמות הללו קשורות למנגנונים שמעורבים בחיסכון במים בתקופה שבה המים הם גורם מגביל, ועוזרים שתישמר בצמח יכולת לביצוע פוטוסינתזה. מרבית המחקרים ברמה הפיסיולוגית אכן עוסקים במאזן תמים ובפוטוסינתזה, אולם העמקת ההבנה של מנגנונים ביוכימיים בצמחים סקלרופיליים עדיין כרוכה בקשיים טכניים ניכרים, בעיקר בגלל הרמה הגבוהה של חומרים משניים בתאים. חומרים אלה גורמים להשקעות חלבונים ולשיבוש הריאקציות הכימיות, ובכך מקשים על המחקר הביוכימי. יתרה מזו, גם ההגדרה של מצבי עקה ברמה התאית היא כשלעצמה בעייתית, בגלל ההבדלים בין תאים שונים ברקמות ואברים שונים, וכן בגלל הבדלים הנובעים מגיל הצמח. התאמות מבניות הקשר בין יובש לבין המבנה הכללי של עצים או שיחים בחורש יכול להיבחן לפי תכונות המופיעות באותו מין צמח לאורכו של מפל לחות אקלימית. השתנות תכונות המבנה הכלליות נבדקה באלון קוקציפרה לאורך מפל גיאוגרפי-אקלימי מצרפת לספרד (1987 ,Leteme and rambal). נמצא, שככל שתנאי האקלים נעשים יובשניים יותר, ישנה ירידה בגובה הממוצע של השיחים המלווה בירידה בפיטומסה (כמות החומר הצמחי [מבוטא ביחידות משקל חומר יבש ליחידת שטח]) העל-אדמתית שלהם. בד בבד חלה גם ירידה באינדכס שטח העלים (leaf area index = LAI). אינדכס שטח העלים הוא היחס בין שטח הפנים הכללי של עלוות צמח לבין שטח ההיטל של נוף הצמח על הקרקע. LAI גבוה מבטא צפיפות גדולה של עלים ושכבתיות מפותחת של ענפים. LAI נמוך קשור, כנראה, בשמירה על מאזן מים תקין בדרך של הפחתת שטח הפנים מאבד המים. ירידת LAI תואמת את התופעה הכללית שבה ערכים אלה גבוהים יותר בעצים ובשיחים של חורש מאשר בשיחים או בני שיח של פריגנה (בתה) (Mooney, 1983 וראה גם בטבלה 6). 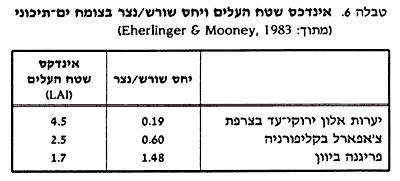 הירידה ב-LAI באלון קוקציפרה עם העלייה ביובש, מקרבת אותו באזורים הקיצוניים לערכים של צמחי בתה. יש לזכור, שערך נמוך של LAI מבטא פוטנציאל נמוך יותר של פוטוסינתזה ברמת הצמח השלם (Mooney, 1983). תכונות מורפולוגיות נוספות של הנצר, שעשויות לסייע בהתגוננות כנגד עקת יובש, הן הופעת עלים קטנים יותר ככל שגדל היובש והימצאות עלים הנטויים בזווית חדה כלפי הענפים (Ehleringer and Mooney, 1983; Miller, 1983). הביטויים האנטומיים של הסקלרופיליות במבנה העלה הם: המצאות תאים קטנים, דפנות תא עבים יותר, צפיפות עורקים גדולה, קוטיקולה (שכבה חיצונית של חומר שומני הקרוי קוטין העוטה את האפידרמיס של הצמחים) עבה יותר ורקמת עמודים מוגדרת היטב ומפותחת. בצמחי חורש ירוקי-עד, השטח הכללי שתופסים נקבי הפיוניות מתוך כלל שטח העלה הוא נמוך (0.5-0.2%) בהשוואה לממוצע של מרבית הצמחים (1.5-0.5%). לעתים, הפיוניות נתונות בתוך שקעים או חריצים המכילים שערות (1981 ,Margaris ;1974 ,Kummerow). כל התכונות האלה מקובלות כתכונות קסרומורפיות, דהיינו, תכונות האמורות להקנות עמידות ליובש. במחקר אנטומי-השוואתי שערך Kummerow (שנת 1973), נמצאה בצמחים סקלרופיליים מאזורים ים-תיכוניים שונים בעולם וריאביליות גדולה בין המינים שנבדקו, בתכונות רבות (מיקום וצפיפות פיוניות, תכונות רקמת העמודים ועוד). יתרה מזו, ישנה גם שונות גבוהה בתוך המינים הנובעת מתנאי בית גידול שונים, כמו ההבדלים בין עלי שמש לעלי צל. יחד עם זאת, שכבת הקוטיקולה העבה יחסית היא תכונה משותפת לעלים סקלרופיליים רבים, וייתכן שזו התרומה האנטומית העיקרית למניעת איבוד מים. ייתכן גם, שפרנכימת העמודים (רקמות בצמח הבנויות מתאים, שלא חלה בהם התמחות מיוחדת) והסקלרנכימה (רקמת תמך בצמחים שעשויה מתאים מעובי דופן, עשירה על פי רוב בליגנין) המפותחות מקנות לעלים מידה מסוימת של הגנה כנגד נזקים שאינם ניתנים לתיקון במהלך של יובש קיצוני. אולם, בסיכומו של דבר, סבור קומרוב שאי אפשר להקיש על העמידות ליובש רק על סמך התכונות האנטומיות בגלל הווריאביליות הרבה שלהן בצמחים הסקלרופיליים השונים. לדעתו, מתאימה לצמחים אלה ההגדרה של "קסרופיטים מתונים". וריאביליות בתכונות אנטומיות של עלים סקלרופיליים נמצאה גם ע"י קריסטודולאקיס ומיטראקוס (1987) בצמחי החורש של יוון. אמנם, עוביה של הרקמה הפוטוסינתטית (המזופיל) בעלים הסקלרופיליים השונים שנבדקו הוא פחות או יותר דומה, וגם האפידרמיס העליון והתחתון מפותחים, אך חלקו של האפידרמיס מתוך סה"כ העובי של העלים משתנה ונע בין 7.4% בזית עד ל- 2.4% בהרדוף. בחלק מהמינים, הדופן החיצוני של תאי האפידרמיס תופס חלק עיקרי בנפח הכללי של התא. בדרך כלל, צפיפות הפיוניות גדולה יותר בעלים הקטנים לעומת הגדולים. מערכת שורשים מעמיקה גם היא תכונת מפתח לשמירה על מאזן מים תקין של צמחים סקלרופיליים בתקופת היובש הקיצי ולהימנעות ממצב עקה (1983 ,Ehleringer and Mooney). תכונה זו באה לידי ביטוי במיוחד בסלעים סדוקים שבהם מי גשמי החורף יכולים לחלחל ולהיאגר בסדקי הסלע. בבתי גידול כאלה נצפו שורשים המגיעים לעומק של 10-5 מטרים ויותר (Walter, 1973; Margaris, 1981). בשנת 1955 ו-1962 Zahary מציין שלמרבית צמחי החורש בארץ, הגדלים על מדרונות סלעיים, יש מערכת שורשים מ"טיפוס הזית". מערכת שורשים כזאת מתפצלת בקירבת צוואר השורש, חלק מהשורשים צומחים בכיוון אופקי, ואילו מרבית השורשים יורדים לעומק העולה על 4 מטרים. השורשים העמוקים הם התורמים לאיזון משק המים בקיץ. מחקר מקיף שנעשה בכרמל בתחילת שנות השישים הביא תוצאות מעניינות על האבפוטרנספירציה (איבוד מים דרך הצמחים) ויעילות ניצול המים של עצי יער וצמחיית חורש (שחורי, 1966; Schori et al., 1966 ). מהמחקר עולה שצמחיית החורש (בעיקר אלון מצוי, אלת מסטיק וסירה קוצנית) הוציאה מים מעומק של כ-8 מטר, משמע מתוך שכבת הסלע הגירני. אורן ירושלים, הגדל על סלעים קירטוניים בשכנות, מוציא מים מעומקים רדודים יותר – עד 5 מ', אך כמות המים הכוללת שהוא מוציא דומה לזו שמוציא החורש. נראה שבקירטון תכולת המים גבוהה בהרבה מזו שבסלעי הגיר והדולומיט. צומח עשבוני משתמש במים הנמצאים בשכבת הקרקע שעומקה 3-1 מ'. ראוי לציין שההבדלים בעומקי הוצאת המים מתייחסים לתקופת היובש, בעוד שבחורף, כל תצורות הצומח מנצלות מים משכבת הקרקע העליונה, שעומקה עד 1 מטר (רוזנצוויג, 1967). את יכולת החדירה של צמחי חורש לתוך שכבת הסלע ניתן להסביר, אולי, בעזרת מנגנוני המסת הסלע על-ידי השורשים כפי שהציע אופנהיימר (Oppenheimer, 1957). פיתוח יתר של מערכת השורשים, כתוצאה מהפניה רבה יחסית של משאבי פחמן לשורש, מבטא, כנראה, עמידות לתנאי צימצום באספקת המים. חלקו של השורש בפיטומסה הכללית של הצמח גבוה במיוחד בצמחי בתה, הגדלים כרגיל בסביבות יותר יובשניות (טבלה 5). אולם, ראוי לזכור שבאלה, מערכת השורשים היא לרוב שטחית ומתפרסת לרוחב (1983, Mooney). רמבל ולטם (1987), מציינים שגם באלון קוקציפרה גדל חלקו היחסי של השורש בקצה היובשני של מפל הלחות, אולם, מעבר לסף מסוים של משקעים, אין הצמחים יכולים לקיים מערכת שורשים מסועפת ומעמיקה דיה, ואז מפתח אלון קוקציפרה מערכת שורשים שטחית, שמגיבה מיידית לכמויות משקעים קטנות. קיומה של מערכת שורשים שטחית בצד המערכת המעמיקה אצל אלון קוקציפרה תורם למאזן המים החיובי שלו בתנאים ים-תיכוניים ומאפשר לו גם יכולת תחרות עם צמחים עשבוניים (Rambal, 1984). למרות האמור עד כה בקשר למערכות שורשים, טוען Kummerow (1983), שקשה להצביע על דגם אחיד של מערכות שורשים של צמחי חורש וצ'אפארל. מערכות השורשים הן מאוד פלאסטיות בפוטנציאל האדפטציה שלהן לטווח רחב של עומקים, בהתאם לנתוני הקרקע והסלע המיוחדים לכל בית גידול. התאמות פיסיולוגיות צמחי החורש הסקלרופיליים שומרים על מופע ירוק כל השנה, ולפיכך יש להם כושר פוטנציאלי לקיים פוטוסינתזה ברציפות ובכך להגיע לפרודוקטיביות גבוהה. שאלת המפתח בהתאמה הפיסיולוגית של הצמחים לתנאי החום והקיץ הים-תיכוניים היא, אם כך, יכולתם לאזן את משק המים בצורה תקינה, ובו בזמן לקיים פוטוסינתזה. ערכים אוסמוטיים ופוטנציאל מים וולטר (1973) השתמש בערכים האוסמוטיים של מוהל התאים כאינדיקציה למצב המים של העלים ("הידרטורה" כלשונו). נמצא, שבצמחי חורש סקלרופיליים התנודות במהלך עונות השנה הן, בדרך כלל, קטנות יחסית. הערכים האוסמוטיים בעונת היובש גבוהים רק ב-4-5 אטמוספירות מעל 21 אטמ' (שהוא הממוצע השנתי). רק בביוטופים קיצוניים מאוד ניתן למדוד ערכים שבין 30 ל-50 אטמוספירות. נתונים מצמחי הארץ שמביאים זהרי ופוליאקוב-מיבר וחובריו (זהרי 1955; 1962 ,zohary; פוליאקוב-מיבר, מאיר וקולר 1959) הם הרבה יותר וריאביליים. בכמה מהמינים, כגון אשחר רחב-עלים, בר-זית בינוני וזית אירופי, עולים הערכים הקיציים עד לסביבות 40 אטמ'. במינים אחרים: ער אציל, קטלב מצוי, אלון מצוי, חרוב מצוי ואלת המסטיק, הערכים נעים בין 20 ל-30 אטמוספירות. פוליאקוב (1945) הראתה שהלחץ האוסמוטי בעלי זית, שקד וחרוב היה תמיד גבוה יותר מכח אחיזת המים של הקרקע. משמע, שהאחרון אינו מהווה גורם מגביל בקליטת מים מהקרקע. יצוין שהערכים האוסמוטיים של כמה נשירי חורף שאינם סקלרופיליים, כמו שקד מצוי, אלה א"י, אשחר א"י ועוזרר קוצני, מצויים בתחום דומה. ספק אם הערכים האוסמוטיים יכולים לשמש מדד אמין ובלעדי למשק מים תקין, או כעדות להתאמה ייחודית של הצמחים הסקלרופיליים. מחקרים פיסיולוגיים מודרניים, שהסתייעו במיכשור חדשני, נערכו בשנות ה-80 בכמה צמחי חורש בפורטוגל (Tenhunen et al., 1987). המדד למצב המים בעלים היה פוטנציאל המים שנמדד בעונות שנה שונות ובמהלכים יממתיים בעונות היובש. בסדרות שונות של מדידות נמצאו הערכים הבאים (טבלה 7):  מטבלה 7 נראה כי ערכי פוטנציאל המים של שני המינים הסקלרופיליים נמוכים בהרבה (בעלי ערך יותר שלילי) בקיץ מאשר באביב. דבר זה עשוי להצביע על עקת מים גוברת בקיץ. יחד עם זאת, נראה שיש הבדל בין האלון והקטלב, כאשר הערכים בקטלב יותר קיצוניים אפילו עם שחר. בכך יש אינדיקציה לעקת מים חריפה יותר במין זה. השאלה היא, עד כמה מצליחים הצמחים לקיים פעילות פיסיולוגית במצב זה. אמת המידה היא חילוף הגזים ושיעורי הפוטוסינתזה המתקיימים בתנאים אלה. חילוף גזים ופוטוסינתזה בצמחים סקלרופיליים ככלל, בצמחים ירוקי העד הסקלרופיליים באזורים הים-תיכוניים, מתקיים כושר של פעילות פוטוסינתטית לאורך כל השנה. עם זאת, השיא בקיבוע הפוטוסינתטי של פחמן חל באביב (Mooney, 1983). בעונת החורף והאביב, שבה המים מצויים בכמות מספקת, מתגלה בדרך כלל מהלך יממתי של טרנספירציה (דיות) המתחילה בבוקר עם פתיחת הפיוניות, מגיעה לשיאה בשעות הצהריים ודועכת שוב לקראת הערב. יחד עם זאת, למרות שהפיוניות פתוחות באמצע היום בחורף, שיעורי הפוטוסינתזה נמוכים מאוד, בגלל טמפרטורת אוויר נמוכה. רק באביב ישנה עלייה גדולה בשיעורי הפוטוסינתזה. דגם כזה מאפיין את מרבית העצים ירוקי העד של החורש הים-תיכוני. במחקר על המהלך היומי והשנתי של טרנספירציה בעצי אורן ירושלים, שנערך ברמת הנדיב, נמדדו ערכים של כ-3 מ"מ ליום בימים בהירים בחורף. רמת הדיות עולה לכדי 7-5 מ"מ ליום באביב ודועכת, תוך זמן קצר ביותר, לכדי 1 מ"מ ביום, בהתאם לזמינות המים בקרקע. כמו כן נמצא כי האורן (ללא תת-היער) מנצל כדי 70% מכמות הגשם היעיל (שילר, מידע אישי). המהלך השנתי של שיעורי הפוטוסינתזה תואם בדרך כלל את אופטימום הטמפרטורה של הפוטוסינתזה, אם כי אין אחידות במידע מהמקורות השונים. לפי Mooney (1981), האופטימום הוא 20 מעלות צלזיוס (מתייחס לצ'אפארל בקליפורניה), בעוד שבכמה צמחים סקלרופיליים שנבדקו בפורטוגל, נמצא שאופטימום הטמפרטורה לפוטוסינתזה הוא 20-25 מעלות צלזיוס, מה שמתאים מאוד לטמפרטורת הצהריים באביב וראשית הקיץ. יוצא דופן הוא קטלב אונדו, שאצלו שיעורי הפוטוסינתזה נטו אינם יורדים גם בחודשי החורף הקרים והוא איננו רגיש לטמפרטורות נמוכות (Beyschlag וחובריו, 1986). בקיץ החם והיבש ישנה ירידה בשיעורי הפוטוסינתזה הנובעת מסגירת פיוניות. סגירת הפיוניות מותנית במשכה של תקופת היובש וביכולת הצמחים השונים להשיג מים מתקופה זו. נוצר מצב שמגביל את הפוטוסינתזה לשימוש בפחמן דו-חמצני מתוך מאגרים פנימיים בלבד, כולל מיחזור מפליטת גזים בנשימה. מחקרים אחדים נעשו מאז שנות ה-30 והלאה בקשר למהלך היומי של פתיחת הפיוניות בחדשי הקיץ בצמחי חורש ים-תיכוניים. עבודות כאלה נעשו באיים בים האדריאטי, בקורסיקה, בפורטוגל ובישראל (Tenhunen et al., 1980, 1981, 1987; Guttenberg + Buhr, 1935; Oppenheimer, 1947, Poljakoff, 1946). בחלק מהמחקרים נבדק המהלך היומי של פתיחת הפיוניות בתנאי שדה, ובאחרים נעשו סימולציות בתנאי חממה. נמצא שבמינים סקלרופיליים רבים בקיץ, הפיוניות פתוחות בבוקר, נסגרות לקראת הצהריים, נפתחות שנית אחר הצהריים ונסגרות בערב. מתקבלת עקומה דו-שיאית טיפוסית שמתארת סגירה של פיוניות בשעות השיא של החום בצהריים (ראה איור 7). מהלך כזה נמצא בצמחים הבאים: זית אירופי, חרוב מצוי, אלון מצוי, ער אציל, בר-זית צר-עלים, אלון קוקציפרה, קטלב אונדו, אלון אילקס ואלון השעם. בצמחים אחרים נמצא מהלך יומי היוצר עקומה חד-שיאית בצורה של "כתף", כלומר פיוניות פתוחות בבוקר ולפני הצהריים, וסגירה של פיוניות מהצהריים והלאה. מהלך כזה מאפיין צמחים כמו אלת המסטיק, קטלב מצוי, וקיסוסית קוצנית. הדגמים הללו מתקיימים גם בתנאי חממה, שבהם ממשיכים להשקות את הצמחים. מכאן מסיקים Lange וחובריו (1982) שמכניזם הבקרה אינו קשור למצב המים בקרקע אלא לטמפרטורה או לשינויי לחות באטמוספירה. ברור מאליו שהמהלך היומי של פתיחת הפיוניות משתקף במהלך יומי דומה של פוטוסינתזה וטרנספירציה (דיות - איבוד מים מהצמח לסביבה). (ראה לדוגמה איור 7)  צימצום הדיות וסגירת הפיוניות במהלך ימי הקיץ הם, איפוא, התאמות (אדפטציה) בצמחים סקלרופיליים המביאים לחיסכון במים, אך כרוכים בירידה בסה"כ הפוטוסינתזה נטו בחדשי הקיץ. המחקרים שנעשו בצמחי החורש בפורטוגל, בשדה ובתנאים מבוקרים, רומזים שעיכוב הפוטוסינתזה בשעות החום הקשות איננו דווקא תוצאה של סגירת הפיוניות אלא ייתכן שהוא נובע מהשפעה ישירה של טמפרטורה גבוהה או עיכוב של אור על הכושר הפוטוסינתטי של העלה, כנראה ברמת המנגנונים הביוכימיים. יחד עם זאת, ישנם הבדלים בהיבט זה בין מינים סקלרופיליים שונים. כך למשל, אלון קוקציפרה מותאם, כנראה, טוב יותר לעקת יובש מאשר קטלב אונדו ואלון השעם, משום שהכושר הפוטוסינתטי שלו ברמה הביוכימית נפגע פחות בזמן סגירת הפיוניות, והוא מסוגל לנצל פחמן דו-חמצני מן המאגרים הפנימיים, גם כאשר פוטנציאל המים מגיע לערכים שליליים בסדר גודל של -30 בר. ייתכן שההסבר לכך הוא שורשיו המעמיקים יותר, המבטיחים לו אספקת מים משופרת יחסית, גם בשיא הקיץ (Tenhunen et al., 1980, 1981, 1985, 1986), שכן אחרת פוטנציאל המים יכול היה להגיע לערכים נמוכים בהרבה. גם מיני הצמחים השליטים בחורש הישראלי מגיבים בצורה שונה ליובש. אלון מצוי מפסיק לחלוטין את הטרנספירציה וסוגר את הפיוניות. בר זית, לעומת זאת, ממשיך בטרנספירציה בקצב שווה תוך כדי שמירה על רמת פתיחה קבועה של הפיוניות. אלה ארצישראלית ממשיכה לדיית כשמתחזק היובש, אך הקצב מועט בגלל סגירה חלקית של הפיוניות (Oppenheimer, 1953). יוצא איפוא, שצמחי החורש הסקלרופיליים מסוגלים אמנם לבצע פוטוסינתזה במשך כל השנה ובכלל זה גם בקיץ, אך אז היא מוגבלת בעיקר לשעות הבוקר המוקדמות, ובחלק מהמקרים גם לשעות אחה"צ המאוחרות. בסך הכל השנתי, לפוטוסינתזה יש מגבלות הן בחורף, עקב הקור, והן בקיץ עקב סגירת הפיוניות ופגיעה בכושר הפוטוסינתטי. היבט נוסף של הקשר בין הכושר הפוטוסינתטי לתנאי הסביבה הוא מסלול הקיבוע של הפחמן. בדרך כלל, באזורים המצטיינים בטמפרטורות גבוהות ובקרינה חזקה נפוץ מאוד מעגל C4, שתוצריו הם בעלי שרשרת פחמן ארוכה מהרגיל. בחורש הים-תיכוני אין צמחי C4 בין המינים הסקלרופיליים, למרות הקרינה החזקה והטמפרטורות הגבוהות של הקיץ. כנראה שהסלקציה לטובת צמחי C4 פועלת בתנאים הללו בשילוב עם שפע מים, דבר שאינו תופש בקיץ הים-תיכוני (Mooney, 1981). שיעור הפוטוסינתיזה המוגבל בצמחים ים-תיכוניים מוסבר בחוסר היכולת לשמור על רמת חלבונים גבוהה, שהיא תנאי לפיזור המוטמעים (Mooney, 1977), זאת בגלל זמינות נמוכה של מינרלים בקרקע. כתוצאה מכך, קצב קליטת פחמן על ידי צמחי חורש ירוקי-עד סקלרופיליים נמוך ביחס לצמחים אחרים (Mooney, 1977). אם משווים את שיעורי הפוטוסינתזה על בסיס של קליטת פחמן דו-חמצני לשטח עלה ביחידה זמן (מבלי להתחשב במשך החיים של העלים), תופסים צמחי החורש הסקלרופיליים עמדת ביניים בהשוואה לצורות חיים אחרות באקוסיסטמות ים-תיכוניות. הסדר של שיעורי הפוטוסינתזה שנמצא בצורות חיים שונות בחורש ירוק העד בצ'ילי הוא כדלקמן (לפי Ehleringer + Mooney, 1983): נשירי קיץ ← חד שנתיים ← עשבים רב-שנתיים ← עצים ושיחים ירוקי-עד ← צמחים רתמיים ← צמחים היגרופיליים ← סוקולנטים.
העצים והשיחים הסקלרופיליים ירוקי-העד הם, כאמור, המזהים הטובים ביותר של אזורי האקלים הים-תיכוניים בעולם. יחד עם זאת, כפי שהראינו בפרקים הקודמים, קשה להגיע למסקנות חד-משמעיות בקשר לערך האדפטיבי של מכלול התכונות שנמנה במינים הסקלרופיליים הללו, לתנאים הספציפיים של האקלים הים-תיכוני, ובמיוחד ליובש הקיץ. כאן המקום לקיים בירור בקשר לנסיבות ההיסטוריות והאבולוציוניות שבהן הופיעו הצמחים הים-תיכוניים הסקלרופיליים. מקובל על הכל, שמאפיינים שונים של האקלים הים-תיכוני, לא הופיעו באזורים שבהם הוא שורר כיום לפני סוף הפלאוקן או ראשית הפלייסטוקן (לפני כמיליון שנה). עדויות פלינולוגיות, שהושגו משכבות קרקע ימית (דרום צרפת), וניתוח אוכלוסיות הפלנקטון שבהן (בעיקר פורמיניפרה), הראו שהשינוי האקלימי המשמעותי ביותר, שחל באזורנו, התרחש לפני 3.2 מיליון שנה. בתקופה זו הוחלף האקלים הטרופי (גשמי קיץ) - בו התפתח יער ירוק-עד, רחב עלים, בעל תכונות סקלרופיליות (דומה ליערות המצויים כיום בסין) - באקלים ים-תיכוני (יובש בקיץ). שינוי זה גרם להחלפת הצומח ולהשתלטות הסוגים: אלון, אלנוס, בר-זית, זית, לוטם ואלה (Suc, 1984). לפי Raven (שנת 1973), התפתחו הפלורות הים-תיכוניות של חצי הכדור הצפוני (בים התיכון ובקליפורניה) מאז הקרטיקון באזור המעבר בין הפלורות של האזור הממוזג מצפון והאזור הטרופי מדרום. המינים הים-תיכוניים עברו סלקציה מתוך מינים טרופיים וממוזגים כאחד והם השתלשלו מחברות הצומח העשירות ששלטו באזורים התת-ארידיים במשך השלישון. לפי אכסלרוד (Axelrod, 1973), פונס (Pons, 1981) ופונס וקזל (Pons + Quezel, 1985), קיימת מאז הקרטיקון, במהלך השלישון ועד לראשית הפלייסטוקן, מגמה מתמדת של התייבשות שקיבלה תאוצה במיוחד באוליגוקן. יחד עם זאת, בכל האקלימים ששררו אז, היו גשמי קיץ ולרוב היתה זו גם עונת הגשם העיקרית. גשמי הקיץ הלכו ופחתו בהדרגה במהלך הניאוגן והאקלים הים-תיכוני הנוכחי, שמאופיין ביובש קיצי מוחלט או יחסי, ושורר למעשה רק מאז ההתייבשות שחלה מאז תקופת הפלייסטוקן המאוחר. הממצאים המאובנים מראים, שמינים סקלרופיליים רבים היו משולבים בחברות הצומח של השלישון יחד עם צורות חיים אחרות, כמו עצים נשירים ורחבי עלים אחרים, אך לא היו דומיננטיים. ככלל, הצמחייה היתה אז עשירה יותר והצומח דמה לזה שמצוי כיום באיים הקנריים ובמדאירה. ככל שהאקלים התייבש, נוצרו בתי גידול צחיחים יותר, במיוחד במדרגות הרום הנמוכות, שהביאו, כנראה, לשיכלול של תכונות סטרוקטוראליות ופונקציונליות כמו: שורשים מעמיקים, עלים יותר ירוקי-עד וסקלרופיליים, שהתאימו ליובש שגבר. בבתי גידול אלה ייתכן שלסקלרופיליים היה יתרון. המאובנים מאירופה, מאגן הים התיכון ומהסהרה מגלים את הצמחים הסקלרופיליים הנוכחיים או את קרוביהם ואבותיהם, לאורך כל הרצף הגיאולוגי מהקרטיקון דרך השלישון. מכאן, שהצמחים הסקלרופיליים הם קדומים יותר מהאקלים הים-תיכוני הנוכחי, שהופיע רק בפלייסטוקן המאוחר. יוצא, אם כך, שכל ההתאמות במבנה ובתיפקוד של הצמחים הסקלרופיליים כלל לא נבררו או עוצבו ע"י האקלים הים-תיכוני הנוכחי. התכונות הללו הן פרה-אדפטציה (תכונה ותיקה שמתגלה כמותאמת לתנאי סביבה חדשים) ליובש הקיץ, שהתפתחה באתרים ששררו בהם וריאציות יותר קיצוניות של התנאים האקלימיים של השלישון, או בבתי גידול פחות נוחים. מאידך גיסא, טוען מאי (1989 ,Mai) שהפלורה הקסרומורפית (יובשנית) הים-תיכונית היא תופעה צעירה ביותר, שהתפתחה רק בפלייסטוקן (אפילו שיש לה שורשים עתיקים יותר) - זאת לפי ממצאי מאובנים שמתארכים את היעלמות תצורות הצומח שקדמו לתצורה הים-תיכונית ואת ראשית הופעת המינים המוכרים כיום. תהליך דומה תואר על-ידי Specht (שנת 1981) לגבי האבולוציה של הצומח הים-תיכוני באוסטרליה. גם כאן נראה שמוצא הפלורה הסקלרופילית הוא בצמחייה טרופית-סובטרופית. אותם מינים שהיו בעלי התאמה ליובש עונתי נבררו בתהליכי סלקציה, כאשר האקלים התייבש במהלך תקופת הרביעון. עדות להתפתחות זו רואה Specht בכך, שתקופת הצימוח הווגטטיבי של מינים ים-תיכוניים רבים בדרום אוסטרליה ודרום אפריקה היא בסוף האביב-קיץ, תקופה שהיא היבשה ביותר באקלים הים-תיכוני הנוכחי אך היתה, קרוב לוודאי, נוחה לצימוח במשטר האקלימי הקודם בו התפתחו הצמחים האלה. שמידע (1980) מדגיש אף הוא את החשיבות של ה"ירושה הפילוגנטית" של צמחיית החורש הים-תיכוני בהבנת מנגנונים פיזיולוגיים או אקולוגיים המאפיינים צמחייה זו על רקע תנאי הסביבה העכשוויים. בניגוד לכך, שימש האקלים הים-תיכוני הצעיר מאיץ להתפתחות אבולוציונית נמרצת ולהיווצרות מוגברת של מינים חדשים (ספציאציה), דווקא של מינים עשבוניים ובמיוחד חד-שנתיים. Pignatti (שנת 1978) מציע מספר גורמי מפתח באבולוציה של הפלורה הים-תיכונית מאז הרביעון: ספציאציה כתוצאה מפתיחת שטחים גדולים (נישות חדשות ←צמחים אנדמים); הפרדה גיאוגרפית בין מינים ממוצא משותף; שינויים במבנה הצמחי (מעוצה ←עשבוני, ירוק-עד ←נשיר קיץ, רב-שנתי ←חד-שנתי; ושינויים גנטיים - פוליפלואידיות, היברידיות) בעיקר כתוצאה מפעילות האדם באלפי השנה האחרונות. התוצאה הסופית היא פלורה עשירה ומגוונת, עם דרגה גבוהה של אנדמיזם. אכסלרוד משער, שאם שוב תחול תנודה והאקלים ישתנה בכיוון של עלייה בכמות המשקעים ותיגבור גשמי הקיץ, יוכלו המינים הסקלרופיליים לשרוד, בעוד שלחד-שנתיים צפויה הכחדה.
בד בבד עם העצים והשיחים הסקלרופיליים הדומיננטיים, גדלים בחורש הים-תיכוני גם שיחים לא סקלרופיליים שמשירים חלק מעלוותם בקיץ. לאלה עלים פחות נוקשים ומשקלם הסגולי של העלים נמוך מזה של העלים הסקלרופיליים. השקעת תרכבות הפחמן בהם מועטה יותר, כלומר ייצורם "זול" יותר מבחינה אנרגטית מזה של עלים סקלרופיליים. מהלך החיים של עלים כאלה, כפי שנחקר במיוחד במינים שונים של הסוג לוטם (Cistus), שונה אף הוא מזה של העלים הסקלרופיליים: משך הזמן שבו נוצרים עלים חדשים הוא ארוך בהרבה. הלבלוב מתחיל בו זמנית עם גשמי הסתיו, והצמיחה נמשכת לאורך החורף והאביב, עד לתחילת הקיץ. מצד שני, תחלופת העלים מהירה יותר מאשר אצל הסקלרופיליים - משך החיים של העלה הבודד הוא בממוצע 5-4 חודשים בלבד (Pereira et al., 1987). בקיץ עומדים השיחים בשלכת חלקית; מרבית העלים שליבלבו בחורף ובאביב נושרים בראשית הקיץ והעלווה הקיצית מקורה בליבלוב המאוחר של ראשית הקיץ. לדוגמה, במין הלוטם Cistus salvifolius, שיעור הנשירה בשנים יבשות עלול להגיע עד 80% (Harley et al., 1987). מערכת השורשים של שיחים לא-סקלרופיליים היא בדרך כלל שטחית יותר מזו של עצים ושיחים סקלרופיליים, אך היחס שורש/נצר הוא גבוה יותר, דבר המצביע על מערכת שורשים מאוד מסועפת, אם כי לא מעמיקה. השרת העלים נחשבת כמנגנון של הימנעות מיובש, בהשוואה לסבילות ליובש שמגלים הצמחים הסקלרופיליים השומרים על עלווה ירוקה במשך כל הקיץ. הישארות חלק מהעלים על הצמח בקיץ מאפשרת לצמחים משירי קיץ חלקיים, כמו מיני הלוטם, לקיים אסטרטגיית התאמה המשלבת סבילות ליובש עם הימנעות מיובש. בכמה מינים (Cistus salvifolius ו- Cistus ladanifer), עלי הקיץ הם יותר נוקשים ובעלי משקל סגולי גבוה יותר, דבר המקרב אותם לסקלרופיליים. ברמה הפיסיולוגית, ניתן להצביע על כמה תכונות המבדילות את התנהגות מיני לוטם מהתנהגות מינים סקלרופיליים:
המידע מבוסס על מחקרים שנעשו ב- 4 מינים של לוטם בפורטוגל (Correeia et al., 1987; Harley et al., 1987) לסיכום, שיחי חורש לא-סקלרופיליים (כדוגמת מיני הלוטם) מותאמים במשק המים שלהם לקיץ החם והיבש, בדרך של איבוד חלק ניכר של העלווה. בכך נפגע אמנם חלק גדול מכושרם הפוטוסינתטי ברמת הצמח השלם, אולם הפיצוי הוא בקיום עונת צמיחה ממושכת יחסית, בפרודוקטיביות גבוהה יותר בחורף ובראשית האביב, בהשוואה לסקלרופיליים, ובהשקעה פחותה של פחמן לבניית עלה נוקשה וצפוף כמו אצל הסקלרופיליים. בסך הכל, המאזן הכולל של הפרודוקטיביות דומה בשני הטיפוסים המורפו-אקולוגיים הללו. א. האקלים הים-תיכוני ב. החורש הים-תיכוני בעולם ג. החורש באגן הים התיכון ד. החורש הים-תיכוני בישראל ה2. התאמות של צמחי החורש לסביבה (חלק 2) ה3. התאמות של צמחי החורש לסביבה (חלק 3) ו1. השפעת האדם על הצומח הים-תיכוני (חלק 1) ו2. השפעת האדם על הצומח הים-תיכוני (חלק 2) ז. ממשק החורש הים-תיכוני
|
|||||||||||||||||||||||||||||||||||||||||||||
 |
|
|||||||||||||||||||||||||||||||||||||||||||||
| 123 |