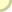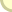|
|
|
   |
הסדרי נגישות
|
| עמוד הבית > מדעים > ביולוגיה |
|
|||||||||||||||||||||||||||||||
|
|
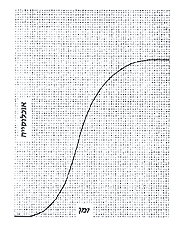
רודריק מארצ'יסון ויתר, בעידודה הנמרץ של אשתו, על התענוגות הכרוכים בציד שועלים והמירם בהנאות היותר נשגבות של המחקר המדעי. הגיאולוג האריסטוקראטי הזה הקדיש חלק גדול מהקאריירה השנייה שלו לתיעוד השלבים הראשונים בתולדות החיים. הוא גילה כי ההתמלאות הראשונה של האוקיאנוסים ביצורים חיים לא התחוללה בהדרגה עם התוספת הרב-שלבית של צורות חיים הולכות ומשתכללות בהתמדה. תחת זאת נראה לו כי מרבית הקבוצות הגדולות נולדו בעת-ובעונה-אחת, במה שהגיאולוגים מכנים כיום הבסיס של תקופת הקאמבריון, לפני כ-600 מיליון שנה. מארצ'יסון, חסיד בריאה מסור, שחיבר את כתביו בשנות השלושים של המאה התשע-עשרה, חשב כי לאירוע הזה יש רק משמעות אחת - אלוהים החליט סוף-סוף לאכלס את כדור-הארץ. צ'ארלס דארווין השקיף על התצפית הזאת בחלחלה. הוא הניח, כמתחייב מן האבולוציה, שהימים "שקקו יצורים חיים" גם לפני תקופת הקאמבריון. בניסיון להסביר את העדרם של המאובנים מן התיעוד הגיאולוגי הקדום, העלה דארווין השערה שיש בה מן ההתנצלות, שיבשותינו המודרניות לא צברו שום משקעים במרוצת הפרקאמבריון לפי שהיו מכוסות אז בימים צלולים. ההשקפה המודרנית שלנו משלבת את שתי ההשקפות הללו. דארווין צדק, כמובן, בטענתו המכרעת: החיים הקאמבריוניים אמנם נולדו מתוך צורות אורגאניות קודמות, לא היישר מידו של האלוהים. אך התצפית היסודית של מארצ'יסון משקפת מציאות ביולוגית, לא פגמים בראיות הגיאולוגיות: תיעוד המאובנים הפרקאמבריוני (פרט לשוליו הקיצוניים) הוא קצת יותר מ-2.5 מיליארד שנים של חיידקים ואצות. החיים המורכבים אמנם נולדו במהירות מדהימה בסמוך לבסיס של הקאמבריון. (על הקוראים לזכור כי לגיאולוגים יש מושגים משלהם בנוגע למהירות. פתילה הדולקת במשך 10 מיליון שנים היא פתילה אטית מאוד על-פי אמות-מידה יומיומיות, אבל 10 מיליון שנים הם רק 1/450 של תולדות כדור-הארץ, הרף-עין לדידו של גיאולוג.) הפאליאונטולוגים השקיעו מאה שנים, עקרות ברובן, בניסיון להסביר את ה"מפץ" הקאמבריוני הזה - העלייה התלולה במגוון במשך 10 או 20 מיליון השנה הראשונות של תקופת הקאמבריון (ראה פרק 14). הם הניחו, פה אחד, שהמאורע המתמיה הוא המפץ עצמו. מכאן עולה שכל תיאוריה משכנעת צריכה להסביר מה עשה את ראשית הקאמבריון לזמן לא-רגיל כל-כך: אפשר שמדובר בהצטברות הראשונה של חמצן אטמוספירי די הצורך לנשימה, או בהתקררות של כדור-הארץ שקודם לכן היה חם מכדי לקיים חיים מורכבים (אצות פשוטות יכולות לשרוד בטמפרטורות גבוהות בהרבה מאלו היפות לבעלי-חיים מורכבים), או בשינוי בכימיה של האוקיאנוס, שהתיר התרבצות של סידן פחמתי וכיסוי של בעלי-חיים רכי-גוף בשלדים בני-שימור. בימים אלה מקננת בי התחושה שהמקצוע שלי צפוי במהרה לשינוי-גישה יסודי. אולי השקפנו על הבעיה החשובה הזאת מן הצד הלא נכון. אולי היה המפץ כשלעצמו רק תוצאה חזויה של תהליך שכבר נכנס למסלול איתן בעקבות מאורע שהתרחש עוד בפרקאמבריון. אם כן הדבר, כי אז לא נצטרך לקבל את ההנחה שימי הקאמבריון היו "מיוחדים" באיזשהו מובן; יהיה עלינו לחפש את הגורם למפץ במאורע מוקדם יותר שפתח את האבולוציה של החיים המורכבים. לאחרונה השתכנעתי כי סביר להניח שנקודת-מבט חדשה זו היא נכונה. מתעורר הרושם כי המתכונת של המפץ הקאמבריוני כפופה לחוק כללי של צמיחה. החוק הזה חוזה שלב של תאוצה תלולה; המפץ עצמו אינו יסודי (או נצרך להסבר מיוחד) יותר מתקופת הגדילה האטית שקדמה לו או ההתאזנות שבאה בעקבותיו. אותו דבר, יהיה אשר יהיה, שפתח את התקופה הקודמת, הוא אשר סיפק למעשה ברבות הימים גם את הערובה למפץ. אני מבקש לתמוך את נקודת-המבט החדשה הזאת בשני טיעונים, המבוססים על כימות של תיעוד המאובנים. הריני מקווה כי יעלה בידי לא רק להוכיח את הנקודה המסוימת האמורה אלא גם לאייר את התפקיד שהכימות יכול למלא בבדיקת השערות בתחומים שנמנעו לפנים מן העוצמה הזאת. עבודת-השדה היומיומית של הגיאולוג היא לכאורה תרגיל מפרך שעיקרו ירידה לפרטי-פרטים: מיפוי של שכבות; חישוב מתאם הזמן שלהן על-פי מאובנים ועל-פי "סופרפוזיציות" פיסיות (שכבות צעירות על שכבות זקנות); תיעוד של סוגי הסלעים, ממדי הגרגירים וסביבות ההתרבצות. תיאורטיקנים צעירים וזריזים מרבים לזלזל בפעילות הזאת ולראות בה עבודה אפורה של חוקרים בינוניים ונטולי-דמיון. ועם זאת, שום מדע לא יקום בלא היסודות שמספקים הנתונים הללו. בהקשר זה, הפרספקטיווה המתוקנת שלנו על המפץ הקאמבריוני מבוססת על שיפור של תיאור השכבות הקאמבריוניות המוקדמות שהוא בעיקרו פרי עבודתם של גיאולוגים סובייטיים בשנים האחרונות. הללו שבו וחילקו את הקאמבריון התחתון הארוך לארבעה שלבים ותיעדו את ההופעות הראשונות של מאובנים קאמבריוניים בדייקנות גדולה בהרבה ממה שנודע קודם לכן. עכשיו יש לאל-ידנו להציג סדר מפורט בהרבה של הופעות ראשונות, בשונה מן התיאורים הקודמים שכל הקבוצות סומנו בהן בתווית "קאמבריון תחתון" (והדגישו בדרך זו את המפץ למראית-עין). ג'.ג'. ספקוסקי, פאליאונטולוג באוניוורסיטת רוצ'סטר, מצא לאחרונה כי העקומה המתארת את גידולו של המגוון האורגאני כנגד הזמן, משלהי הפרקאמבריון עד סוף ה"מפץ", עולה בקנה אחד עם מודל הצמיחה הכללי ביותר שלנו - מה שקרוי עקומה סיגמואידית1 (דמוית-S). הבה נתבונן בצמיחה של מושבת חיידקים טיפוסית על מצע שלא היה מיושב לפני כן: כל תא מתחלק אחת לעשרים דקות ויוצר שני תאי-בת. האוכלוסייה גדלה בתחילה בקצב אטי. (המקצבים של חלוקת התא הם מהירים כתמיד, אך מספרם של התאים המייסדים הוא נמוך, והאוכלוסייה נבנית אט-אט לקראת תקופת המפץ.) שלב ה"פיגור" הזה יוצר את החלק ההתחלתי של העקומה הסיגמואידית, העולה לאטה. אחר-כך בא שלב המפץ או השלב הלוגאריתמי: כל תא הנמנה עם האוכלוסייה הגדולה מוליד מדי עשרים דקות שני תאי-בת פורים. ברור בעליל שהתהליך הזה אינו יכול להימשך עד אינסוף: "התפוצצות אוכלוסין" לא-מדי-רחוקה תמלא את היקום כולו בחיידקים. בסופו של דבר, המושבה מבטיחה את יציבותה (או את מותה) בכך שהיא תופסת את כל המרחב שלה, ממצה את חומרי המזון שלה עד תום, משחיתה את קנה בחומרי פסולת, וכיוצא באלה. ההתאזנות הזאת קובעת תקרה לשלב הלוגאריתמי שבעקומה ומשלימה את ה-S של ההתפלגות הסיגמואידית. מרחק רב מפריד בין החיידקים לבין האבולוציה של החיים, אולם צמיחה סיגמואידית היא תכונה כללית של מערכות מסוימות, ונראה כי ההשוואה אכן תקפה בהקשר זה. במקום חלוקת תא יש לקרוא התפצלות למינים; במקום מצע אגר של צלוחית מעבדה יש לקרוא אוקיאנוסים. שלב הפיגור של החיים הוא ההתפתחות ההתחלתית האטית של שלהי הפרקאמבריון (כיום מוכרת לנו פאונה צנועה משלהי הפרקאמבריון - בעיקר נבובים [מדוזות ואלמוגים רכים] ותולעים). המפץ המפורסם של תקופת הקאמבריון אינו אלא השלב הלוגאריתמי של התהליך הרציף הזה, ואילו ההתאזנות לאחר תקופת הקאמבריון מייצגת את המילוי ההתחלתי של הגומחות האקולוגיות באוקיאנוסים של העולם (חיי היבשה התפתחו כעבור זמן). אם ההתגוונות הקדומה של החיים התחוללה על-פי חוקי הצמיחה הסיגמואידיים, כי אז אין שום דבר מיוחד במפץ הקאמבריוני. אין הוא אלא השלב הלוגאריתמי של תהליך הנקבע על-פי שני גורמים: (1) המאורע שהתחיל את שלב הפיגור עוד בעיצומו של הפרקאמבריון ו-(2) התכונות של אותה סביבה שהתירה צמיחה סיגמואידית.
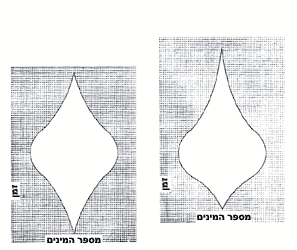
עקומה סיגמואידית (דמוית-S) טיפוסית. ראוי לתת את הדעת ס.מ. סטנלי, הפאליאונטולוג מאוניוורסיטת ג'ון הופקינס, כתב בסקירה שפורסמה לאחרונה (1976, American Journal of Science): "יכולים אנו לנטוש את ההשקפה המסורתית שמוצאן של קבוצות מאובנים גדולות בסמוך לראשית הקאמבריון... מציג חידה גדולה. מה שנשאר כ'בעיית הקאמבריון' היא העובדה שבעלי-החיים הרב-תאיים הראשונים הופיעו לראשונה רק בעת שכדור-הארץ היה בן 4 מיליארד שנים כמעט." נוכל לכפור בבעיית הקאמבריון אם נטילה לאחור, על אירוע מוקדם יותר, אבל צביונו ומחוללו של האירוע הקדמוני הזה עודם חידת-החידות הפאליאונטולוגיות. ראשיתו של התא האיאוקאריוטי בשלהי הפרקאמבריון צריכה לספק גורם קובע חשוב ביותר. (בפרק 13 אני טוען שרבייה מינית יעילה זקוקה לתא איאוקאריוטי בעל כרומוזומים מוגדרים, ושאורגאניזמים מורכבים לא יכלו להתפתח בלי הרבגוניות הגנטית שמספקת הרבייה המינית.) אך אין לנו מושג, ולוא גם הקל שבקלים, על סיבת היוולדו של התא האיאוקאריוטי - כאשר קרה הדבר סוף-סוף, כחלוף שני מיליארד שנים ויותר מאז האבולוציה של אבותיו הפרוקאריוטיים. במאמר 14 הצעתי את תיאוריית ה"קציר" של סטנלי כדי להסביר מה עורר את העלייה הסיגמואידית בעקבות האבולוציה של התאים האיאוקאריוטיים. סטנלי טוען כי האצות הפרוקאריוטיות הפרקאמבריוניות תפסו את כל המרחב הזמין במקום-חיותן הפוטנציאלי, לא הניחו דריסת-רגל לשום מתחרה ומנעו בכך אבולוציה של כל דבר מורכב יותר. אוכל-הצמחים האיאוקאריוטי הראשון מילא את כרסו בסעודה כלל-עולמית שופעת, אם כי חדגונית, ושיחרר תוך כדי כך מרחב גדול די הצורך לאבולוציה של מתחרים. אפשר להציע ניחושים מרתקים, אך נבצר מאתנו לומר הרבה דברים מעשיים על הגורם הראשון שלי: המחולל שפתח את העלייה הסיגמואידית. על-כל-פנים, מצבנו טוב יותר ביחס לגורם השני - טבעה של הסביבה שהתירה את העלייה הזאת. צמיחה סיגמואידית איננה תכונה אוניוורסאלית של מערכות טבעיות; היא מתרחשת רק בסביבה מסוג אחד. חיידקי המעבדה שלנו לא היו יכולים לגדול בעקומה דמוית-S, אילו קדמה להם אוכלוסייה צפופה בקרקע-המזון, או אילולא נמצא מזון במצע הזה. מתכונות סיגמואידיות מתרחשות רק במערכות פתוחות לא-מוגבלות, במקום שמזון ומרחב נפוצים כל-כך והאורגאניזמים יכולים להתרבות עד שמספריהם עצמם מונעים התרבות נוספת. ברור בעליל שהאוקיאנוסים של ימי הפרקאמבריון יצרו מערכת אקולוגית "ריקה" כזאת - מרחב בשפע, מזון לרוב, ללא התחרות. (האיאוקאריוטים הקדמונים חבו תודה לאבותיהם הפרוקאריוטיים לא רק בעד אספקת מזון מיידית אלא גם בעד שירות קודם בשחרור חמצן לאטמוספירה בתהליך הפוטוסינתיזה.) העקומה הסיגמואידית - עם המפץ הקאמבריוני בשלב הלוגאריתמי שלה - מייצגת את המילוי הראשון של האוקיאנוסים בעולמנו, מתכונת חזויה של אבולוציה במערכות אקולוגיות פתוחות. סביר להניח כי בעלי-חיים המתפתחים במרוצת השלב הלוגאריתמי צריכים להראות מתכונות אבולוציוניות שונות מאלו העולות ברבות הימים במשטר של שיווי-משקל מווסת-עצמו. חלק גדול ממחקרי בשנתיים האחרונות הוקדש להגדרת ההבדלים הללו. עמיתי (ת.ג'.מ. שוף מאוניוורסיטת שיקאגו, ד.מ. רופ וג'.ג'. ספקוסקי מאיניוורסיטת רוצ'סטר, וד.ס. סימברלוף מן האוניוורסיטה של מדינת פלורידה) ואנוכי התקנו מודלים של אילנות-יוחסין אבולוציוניים בתהליך אקראי. לאחר ש"גידלנו" עץ, המשכנו וחילקנו אותו ל"ענפיו" הראשיים ובדקנו את תולדותיו של כל ענף. במרוצת השנים, המשכנו ותיארנו כל "ענף" במה שקרוי דיאגראמת כישור. וזו דרך בנייתה של דיאגראמת כישור: מונים בפשטות את מספר המינים החיים בכל פרק זמן ומשנים את רוחב הדיאגראמה על-פי המספר הזה. אחר-כך מדדנו תכונות אחדות של הדיאגראמות הללו. מדידה אחת, הנקראת C.G., מגדירה את מקומו של מרכז-הכבידה2 (הרי זה, במקורב, המקום שבו הענף רחב ביותר, או מגוון ביותר). אם המקום הזה של המגוון המרבי מופיע בנקודת האמצע של משך הענף, אנו מעניקים ל-C.G. ערך של 0.5 (בחצי הדרך של הקיום הטוטאלי של ה"ענף"). אם ה"ענף" מגיע לשיא המגוון שלו לפני נקודת האמצע - ערך C.G. קטן מ-0.5. בשיטה האקראית שלנו, ה-C.G. הוא תמיד ליד 0.5 - ה"ענף" האידיאלי הוא יהלום הרחב ביותר במרכזו. אבל העולם האקראי שלנו הוא עולם של שיווי-משקל מושלם ואין מתירים בו שום שלבים לוגאריתמיים של צמיחה סיגמואידית; מספר המינים נשמר בקביעות במרוצת הזמן, ושיעורי הכיליון תואמים את שיעורי הבריאה של מינים חדשים. עשיתי חלק הגון משנת 1975 בספירת סוגים מאובנים ורישום משך חייהם, בניסיון לבנות דיאגראמות כישור ל"ענפים" אמיתיים. כיום יש בידי יותר מ-400 "ענפים" לקבוצות שהופיעו ונכחדו ברבות השלב הלוגאריתמי של המפץ הקאמבריוני. ערכן הממוצע עומד על 0.4993 - אי-אפשר לבקש משהו קרוב יותר ל-0.5 של עולמנו האידיאלי בנקודת שיווי-משקל. כן יש תחת ידי מספר דומה של דיאגראמות כישור ל"ענפים" שנולדו במרוצת השלב הלוגאריתמי ונכחדו אחר-כך. ה-C.G. הממוצע שלהם נמוך בשיעור משמעותי מ-0.5. הם מנציחים עולם לא-טיפוסי של מגוון גדל והולך וערכיהם יכולים לשמש כדי לאמוד הן את העיתוי הן את העוצמה של השלב הלוגאריתמי הקאמבריוני. ערכיהם נמוכים מ-0.5 מפני שהם נולדו במשך תקופות של התגוונות מהירה, אך נכחדו בזמנים יציבים של בריאה וכיליון אטיים יותר. הם הגיעו למגוון מרבי בשלב מוקדם בתולדותיהם מכיוון שנציגיהם הראשונים השתתפו בשלב לוגאריתמי של גדילה לא מחסנת, אבל הם נעלמו בקצב אטי יותר בעולם המיוצב שבא לאחר מכן.
דיאגרמות כישור. "מרכז-הכבידה" של הדיאגראמה משמאל הוא 0.5 גישה כמותית מסייעת לנו להבין את המפץ הקאמבריוני בשתי דרכים. ראשית, אנו יכולים לזהות את צביון הגדילה הסיגמואידי שלו ולזהות את מחוללו כמאורע קדום יותר; הבעיה הקאמבריונית נעלמת. שנית, אנו יכולים לחקור את הסטאטיסטיקה של דיאגראמות כישור כדי להגדיר את הזמן והעוצמה של השלב הלוגאריתמי הקאמבריוני. התוצאה המרשימה ביותר של התרגול הזה איננה, לדעתי, ה-C.G. הנמוך מתקופת הקאמבריון, אלא ההתאמה של C.G. מאוחרים יותר למודל אידיאלי של עולם בשיווי-משקל. הייתכן שהמגוון של החיים הימיים נשאר בשיווי-משקל במרוצת כל התמורות שעברו על כדור-ארץ השרוי בתנועה, כל הכליונות ההמוניים, ההתנגשויות של יבשות, הבליעה והיצירה של אוקיאנוסים? השלב הלוגאריתמי של הקאמבריון הכיל את האוקיאנוסים של העולם. מאז ועד היום הפיקה האבולוציה אינסוף ואריאציות מקבוצה מוגבלת של תוכניות-יסוד. חיי הים פיתחו מגוון עצום, הסתגלות רבת-תושייה וגם, אם תורשה לי הערה אנתרופוצנטרית, יופי מופלא. אפשר גם לומר - במידה רבה של חשיבות - שהאבולוציה שהתחוללה מאז הקאמבריון רק מיחזרה את התוצרים הבסיסיים של שלב המפץ. 1. באנגלית Sigmoid Fraud, צירוף שיש בו כדי להזכיר את זיגמונד פרויד (המתרגמת). קראו עוד: מתכונות והטעמות בתולדות החיים : הפנטאגון של החיים
|
|||||||||||||||||||||||||||||||
 |
|
|||||||||||||||||||||||||||||||
| 123 |