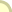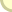|
|
|
   |
הסדרי נגישות
|
| עמוד הבית > מדעים > ביולוגיה > גנטיקה |
|
|||||||||||||||||||||||||||||||
|
|
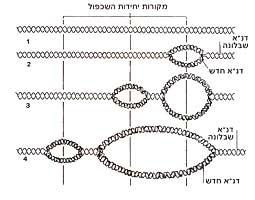
כאן יש כמה שאלות משנה. הראשונה, אולי, היא "כיצד משכפל הדנ"א את עצמו?" השנייה: "כיצד מתחברות שרשראות החלבונים הלכה למעשה, על-פי הוראות הדנ"א?" והשלישית כבר הוצגה לעיל: "איך יכולים ארבעת הנוקליאוטידים השונים בדנ"א לספק קוד מתאים לעשרים סוגי המרכיבים בחלבון? " בדיעבד ניתן לראות כי פולינג ודלבריק כבר ניחשו את עקרון שכפול הדנ"א, וכי ווטסון וקריק רמזו עליו בשורה האחרונה של מאמרם ב-Nature. בפועל, גדיל סלילוני אחד של הדנ"א מתפצל מעל משנהו, ומרגע שמתחיל הפיצול, מתחילים להיערך נוקליאוטידים, המצויים בחומר שמסביב, מול משלימיהם שבגדיל. תוך כדי כך הם מתחברים זה לזה בעזרת האנזים דנ"א-פולימרז (תרשים 2.3). שימו לב - יש לנקודה זו חשיבות מכרעת בהתפתחות ההנדסה הגנטית - עד כמה תלויים פיצול הדנ"א והרכבתו מחדש בפעולתם של צוותי אנזימים, שדנ"א-פולימרז הוא רק אחד מהם.
תרשים 2.3 : שכפול דנ"א העובדה שהדנ"א אכן משתכפל בדרך זו הוכחה לראשונה הלכה למעשה בניסוי מבריק מאין כמוהו שערכו מתיו מיזלסון ופרנקלין סטאל בקלטק ב-1958. הם גידלו תרביות של E.coli בתווך שהועשר בריכוזים גבוהים של האיזוטופים הכבדים 13C ו- 15N. (לאיזוטופים השכיחים של פחמן וחנקן משקלים אטומיים נמוכים יותר, 12 ו-14 בהתאמה, כך שמולקולת דנ"א שנבנתה מן הצורות הכבדות יותר תשקול הרבה יותר.) אז העבירו מיזלסון וסטאל את חיידקי E.coli בעלי הדנ"א הכבד לתווך שהכיל רק את האיזוטופים הקלים יותר של C ו-N , והניחו להם להתחלק פעם אחת. אחרי דור אחד הם מצאו רק סוג אחד של דנ"א בתאי E.coli החדשים, ומשקלו היה באמצע הדרך בין זה של E.coli בעל הדנ"א הכבד לבין E.coli הרגיל, בעל הדנ"א הקל. מכאן היה ברור כשמש שהדנ"א החדש שנוצר הכיל גדיל אחד של דנ"א כבד, וגדיל אחד של דנ"א קל. ברור איפוא שהשכפול התבצע תחילה על-ידי פיצול הדנ"א המקורי הדו-גדילי לשני גדילים נפרדים, ובעקבות זאת שוכפל כל אחד מהם. (העובדה שהשכפול מתחיל מיד כשמתחיל הפיצול, ואינו ממתין עד שתושלם ההפרדה, הוכחה מאוחר יותר.) נעבור איפוא לשאלה הבאה: כיצד יוצר הדנ"א חלבונים? היה ברור מלכתחילה שהדנ"א עצמו אינו מסוגל לפעול כיצרן חלבונים. הדנ"א מוגבל (ברובו) לכרומוזומים שבתוך הגרעין; אבל אפשר להיווכח שחלבונים מסונתזים בציטופלסמה התאית, מחוץ לגרעין. מכאן שחייב להימצא איזה מתווך, המסיע את הוראות הדנ"א לאתרי הייצור של החלבונים. כאן נכנסת לתמונה חומצת הגרעין השנייה, רנ"א. היה סביר לחשוב שאכן הרנ"א הוא המתווך בין הדנ"א שבגרעין לבין אתרי ייצור החלבונים בציטופלסמה, עוד לפני שהציעו ווטסון וקריק את דגם הדנ"א שלהם. למשל, תאים המייצרים חלבונים רבים, תמיד יש שפע של רנ"א בציטופלסמה שלהם; ו"עמודי השדרה" העשויים סוכר וזרחה של הדנ"א והרנ"א דומים מאוד, מה שמצביע על זיקה אפשרית ביניהם. (אם כי בפועל, כפי שראינו, הרנ"א שונה מהדנ"א לא רק בסוג הסוכר - ריבוזה במקום דאוקסיריבוזה - אלא גם בנוקליאוטידים שלו, אף שהשוני קל: הוא מכיל אורציל במקום תימין. עם זאת האורציל יוצר "זוג בסיסים" עם האדנין, בדיוק כמו התימין.) כבר ב-1953, אם כן, החלה להצטייר התמונה: גדיל של דנ"א, לאחר שהופרד משותפו, יכול לעשות אחת משתיים: לבנות העתק משלים של עצמו, וכך להשתכפל, או להתחיל לבנות גדיל משלים של רנ"א, וזה יוצא מהגרעין, עם השלמתו, ומפקח על ייצור החלבון המתאים בציטופלסמה. יצירת הרנ"א המשלים לקטע של דנ"א נקראת שעתוק (טרנסקריפציה). בציטופלסמה, הקוד שנושא עתה הרנ"א הזה בא על ביטויו בחלבון בתהליך הקרוי תרגום (טרנסלציה). בפועל, ייצור החלבון הוא עניין מסובך במקצת. מעורבים בו שלושה סוגים שונים של רנ"א, שכל אחד מהם נוצר על-ידי חלקים שונים של הדנ"א. הרנ"א הנושא את המסר אל מחוץ לתא נקרא רנ"א-שליח. אבל הרנ"א-השליח אינו יוצר את החלבון במישרין. תחת זאת הוא פועל במשותף עם רנ"א השוכן דרך קבע בציטופלסמה (אם כי הוא נוצר מלכתחילה בגרעין), באיברונים הקרויים ריבוזומים, ולכן הוא נקרא רנ"א-ריבוזומי. נוסף על כך, כל חומצה אמינית המיועדת להכללה בחלבון מובלת אל תוך הריבוזום על-ידי קטע קצר של רנ"א הקרוי רנ"א-מוליך. כל רנ"א-מוליך מלווה חומצה אמינית ספציפית ומתקשר למקום מסוים ברנ"א-השליח, כשהרנ"א-הריבוזומי משמש כמין שולחן עבודה שעליו מבוצע החיבור. שוב יש צורך באנזימים כדי לקדם את התהליך בדרכו. הדבר מתואר בתרשים 2.4, אבל למעשה, אין צורך ביותר מאשר התיאור דלעיל.
תרשים 2.4: דנ"א יוצר רנ"א היוצר חלבון הסוגים השונים של רנ"א אינם אלא המוציאים לפועל. ההנהלה המרכזית, הקובעת אילו סוגי חלבונים ייוצרו, היא הדנ"א. מכאן שהשאלה השלישית חייבת להיות זו: כיצד יכול קטע מסוים של דנ"א - גן - לקודד חלבון מסוים? עובדה אחת האומרת דרשני היא שכל חלבון עשוי משרשראות ליניאריות של חומצות אמיניות (גם אם צורתו הסופית של החלבון אינה דומה כלל ועיקר לשרשרת ליניארית), ואילו הדנ"א עשוי משרשראות ליניאריות של נוקליאוטידים. איכשהו, רצף הנוקליאוטידים חייב לקבוע את רצף החומצות האמיניות. אבל כאן אנו חוזרים לשאלה שכבר הוצגה: יש בחלבון עשרים חומצות אמיניות, פחות או יותר, בעוד שבדנ"א יש רק ארבעה סוגי נוקליאוטידים. אם כן, כיצד יכולים ארבעה לקודד עשרים? צירופים - זוהי כמובן התשובה. אם הנוקליאוטידים הערוכים זה לצד זה פועלים בזוגות, אזי ארבעה סוגים שונים יכולים ליצור שישה-עשר צירופים שונים: 4 4x. אם בשלשות, הם יכולים ליצור שישים וארבעה צירופים שונים: 4 4x 4 x, ואם הם פועלים ברביעיות, הם יכולים ליצור 256 צירופים שונים: 4x 4 x 4 x 4. ברור שאין הם יכולים לפעול בזוגות, שכן שישה-עשר זה מעט מדי. מאתיים חמישים ושישה הוא מספר הרבה יותר מדי גדול. קרוב לוודאי, אם כן, שהם פועלים בקבוצות של שלושה. שישים וארבעה הוא מספר גדול מן הנחוץ, ככל הנראה; אבל אין הוא מופרז עד כדי אבסורד. סידני ברנר ופרנסיס קריק, בעבודתם בקיימברידג' ב- 1961, הם שהראו כי הקוד הגנטי אכן פועל באמצעות שלשות של נוקליאוטידים בדנ"א. הם עבדו עם צורות מוטנטיות של הפאג T4, הנטפל לחיידקי E.coli. הם מצאו כי אם מוסיפים רק נוקליאוטיד אחד לגן T4, החלבון המתקבל איננו תפקודי. אם מוסיפים שני נוקליאוטידים, שוב מתקבלים חלבונים לא-תפקודיים. אבל אם מוסיפים שלושה נוקליאוטידים לגן T4, החלבון המתקבל הוא תפקודי לכל דבר. הוספת נוקליאוטיד אחד או שניים רק שיבשה את קריאת הקוד, בלבלה את רצף הקריאה. אבל הוספת שלושה נוקליאוטידים הוסיפה חומצה אמינית שלמה, ששינתה במקצת את החלבון שהתקבל, אבל לא במידה שגרמה לשיבוש רציני בפעולתו. כל שלשה של נוקליאוטידים המקודדת חומצה אמינית מסוימת נקראת כיום קודון. עבודתם של ברנר וקריק גם המחישה בעליל את טיבה של המוטציה. אם גן פועל על-ידי הספקת שלשות של נוקליאוטידים בזו אחר זו - ואכן כך הוא פועל - ברור מאליו שמוטציה יכולה להתרחש רק באחת משלוש צורות אפשריות. אם נוסף לרצף רק נוקליאוטיד אחד, הדבר ישבש קשות את קריאת הקוד. אם נגרע נוקליאוטיד אחד, שוב ישתבש הקוד. אבל אם רק השתנה נוקליאוטיד אחד - כלומר, אם הוחלף נוקליאוטיד אחד במשנהו – הדבר עשוי לגרום (או לא לגרום) להחלפתה של חומצה אמינית אחת בחלבון המתאים, ודבר זה עשוי לשבש (או לא לשבש) את תפקודו של אותו חלבון. מכל מקום, אנו רואים שבמונחים כימיים, מוטציות יכולות להיות שינויים פעוטים, אבל לשינויים פעוטים יכולות להיות תוצאות מרחיקות לכת. בתחילת שנות ה-60 הלכה והתבררה הזיקה בין שלשות מסוימות של נוקליאוטידים לבין החומצות האמיניות, החל בעבודתם של מרשל נירנברג והיינריך מתאי; הם הראו שרנ"א-שליח מלאכותי המכיל אך ורק שרשראות של אורציל יוצר שרשראות של החומצה האמינית פנילאלנין. מכאן ש-UUU, מקודד פנילאלנין. בעקבות זאת נערכו ניסויים דומים במעבדות אחרות, וביוני 1966 כבר היה ידוע שכל הקודונים בנויים משלושה נוקליאוטידים רצופים, וש-61 מתוך 64 הקודונים מקודדים חומצות אמיניות (ברור איפוא שלכמה חומצות אמיניות מתאים יותר מקודון אחד), ואילו השלושה הנותרים ( UAG ,UAA, ו-UGA) משמשים כ"תמרורי עצור" המציינים, פשוטו כמשמעו, שבניית החלבון צריכה להיפסק. בקיצור, הקוד הגנטי "פוצח". כך קלטה הגנטיקה את התחום הנוסף, ביולוגיה מולקולרית, וכך נולדה "הגנטיקה המולקולרית" כתחום מחקר בפני עצמו. במשך שאר הספר הזה נדבר לפעמים על הגנים כאילו אין הם אלא הפשטות בעלמא, כמו חרוזים בשרשרת, ולפעמים נדון בהם כיישויות כימיות ספציפיות שאפשר לפעול עליהן בדרכים ספציפיות. אין להתפלא על כך שביולוגים מולקולריים רבים חשו זחיחות דעת רבה באמצע שנות ה-60. מבנה הגן נפתר, ודומה היה שגם אופן פעולתו נחשף בפירוט רב למדי. שאלה גדולה אחת שעדיין לא נגענו בה (והיא נושא דיונו של הפרק הבא) פוצחה אף היא: דהיינו, כיצד מתחילים הגנים לפעול וכיצד הם מפסיקים לפעול, במרוצת חייו של האורגניזם. כשאוחדה הביולוגיה המולקולרית עם הגנטיקה המנדלית, וכשאוחדה הגנטיקה המנדלית עם רעיונות דארווין על האבולוציה שתוארו בפרק 1 - ובכן, התוצאה נראתה אדירה באמת ובתמים. למען האמת, דומה היה שאין מקום לשפר אותה. כל שנותר, חשבו אחדים, הוא רק ללטש את פרטי הפרטים האחרונים. פרנסיס קריק עצמו ביקש לו אתגרים חדשים בתחום שונה לגמרי: חקר העצבים. לא היה יסוד לזחיחות הדעת הזאת. קשה מאוד לרדת לחקרו של הטבע. שלושים השנים האחרונות הניבו - ומוסיפות להניב - גילויים שאינם סתם קישוטים, אלא הם בסיסיים מאוד: אדירים במידה כזו, בכמה מקרים, שיש בהם כדי להעלות את הביולוגיה כולה על פסים חדשים. נעיין בקצרה באחדים מהגילויים הללו בעמודים הבאים, ונבדוק כמה מהשלכותיהם בפרקים הבאים. לפריטים נוספים מתוך הפרק: הגן הלכה למעשה
|
|||||||||||||||||||||||||||||||
 |
|
|||||||||||||||||||||||||||||||
| 123 |